Morfološki, citološki i fiziološki sterilitet masline
Maslinarstvo
03.04.2013.
Morfološki, citološki i fiziološki sterilitet masline
Urod maslina
Foto: Gemini 2.5 Flash
Slabi urod naših sorata maslina i loše zametanje plodova, iako na stablu ima dovoljno cvjetova, dva su momenta zbog kojih treba pisati i govoriti o toj temi.
Autori, koji su proučavali pojavu morfološkog i fiziološkog steriliteta sorata masline, imaju različita, a nekad i kontroverzna mišljenja. Odgovor, zašto je to tako, možda leži u tome što su radili u različitim ekološkim prilikama i s različitim sortama masline. Naprotiv, citogenetička, a naročito citokemijska proučavanja sorata masline, vrlo su oskudna ili sasvim nedostaju.
U desetogodišnjem istraživanju prve rezultate i analize donosi dr. sc. Aleksandar Vlašić iz Splita.
U svom radu, u svrhu utvrđivanja intenziteta pojave i uzroka lošeg uroda sorata masline, analizira tri osnovna problema:
intenzitet i uzroke pojave funkcionalno muških cvjetova masline;
uzroke niske klijavosti polena sorata masline i
autokompatibilitet i interkompatibilitet sorata masline.
Kad je riječ o uzrocima morfološke sterilnosti cvjetova sorata masline, mišljenja različitih autora su oprečna. Dok jedni tvrde da je taj oblik sterilnosti sortna, odnosno konstantna karakteristika, koju vanjski čimbenici ne mogu mijenjati, drugi smatraju da je morfološka sterilnost uvjetovana isključivo ekološkim čimbenicima. Treća grupa autora zaključuje da su i genetski i ekološki čimbenici uzroci intenziteta morfološke sterilnosti.
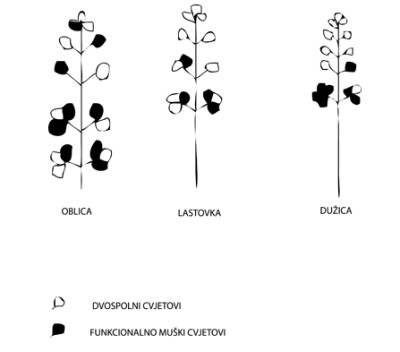
Izneseni rezultati dr. Vlašića o tome kod ispitanih sorata masline pokazuju, prije svega, da je morfološka građa cvata sortna karakteristika. Način grupiranja cvjetova u cvatu, kao i postotna zastupljenost pojedinih cvatova, ovisi o sorti. U tom pogledu ispitane sorte lastovka, oblica i dužica, međusobno se razlikuju. Skladno tome je i ukupan broj cvjetova u cvatu, koji kod lastovke iznosi 14, kod oblice 18, a kod dužice 17.
Međutim, morfološka građa cvata varira od godine do godine pa, prema tome, ne predstavlja nepromijenjenu veličinu, kao što tvrde neki autori. Isto tako varira i broj cvjetova u cvatu od godine do godine. Variranje broja cvjetova u cvatu najveće je kod dužice (11-25), nešto manje je kod oblice (15-23), dok je najmanje kod lastovke (12-17). Osjetno variranje u morfološkoj građi cvata od godine do godine ovisi, pored ostalog, kako je utvrdio dr. Vlašić analizom, o intenzitetu uroda stabla u prethodnoj godini. Obično nakon godine s velikim brojem cvjetova slijedi godina s manjim brojem cvjetova u cvatu.
Mnogo interesantnija su proučavanja stanja hermafroditnih i funkcionalno muških cvjetova u cvatu. Broj hermafroditnih cvjetova u cvatu sortna je karakteristika. Višegodišnji prosjek pokazuje da lastovka, od ukupno 14 cvjetova u cvatu, ima 10 hermafroditnih ili 71,5 %, oblica od 18 cvjetova u cvatu ima svega 9,5 hermafroditnih ili 51,5 %, a dužica od 17 cvjetova ima 9 hermafroditnih ili 54 %. Prema tome, kod oblice se 48,5 %, a kod dužice 46 % cvjetova na stablu ne mogu oploditi. Taj je postotak kod lastovke znatno manji i iznosi 28,5 %.
Slika 1. Cvat oblice, lastovke i dužice s prosječnim brojem hermafroditnih i funcionalno muških cvjetova
Razumljivo je da se tako veliki broj funkcionalno muških cvjetova na stablu kod oblice i dužice mora negativno odraziti na rod. Zato se to istraživanje ne slaže s mišljenjima nekih autora, koji tvrde da se pojava muških cvjetova može zanemariti. Rezultati proučavanja te pojave idu u prilog tvrdnji, koja ističe da je pojava funkcionalno muških cvjetova često uzrok nerodnosti sorata masline. Stoga treba istaknuti da je neredoviti urod kod oblice i dužice često posljedica visokog postotka funkcionalno muških cvjetova. Lastovka ima znatno manji broj funkcionalno muških cvjetova, zbog toga ona redovitije rađa.
Hermafroditni odnosno funkcionalno muški cvjetovi imaju svoj određeni položaj na osi cvata, koji je različit ovisno o sorti. Dakle, položaj hermafroditnih cvjetova u cvatu sortna je karakteristika.
Navedene prosječne vrijednosti za broj funkcionalnih muških cvjetova rezultat su višegodišnjih proučavanja. One su različite za pojedinu sortu i rezultat su sortnih osobina maslina na određenom staništu.
Međutim, rezultati proučavanja dr. Vlašića govore da pojava funkcionalno muških cvjetova nije konstantna i nepromjenjiva veličina. Velika varijacija u intenzitetu ispitane pojave sterilnosti u pojedinim godinama svjedoči da je ta sortna osobina vrlo varijabilna veličina i da je ona pod utjecajem djelovanja i drugih čimbenika. Taj rezultat je u suprotnosti s tvrdnjom nekih autora, po kojima je morfološka sterilnost cvjetova stalna, nasljedna, nepromjenjiva sortna osobina, koju vanjski čimbenici ne mogu mijenjati.
Iz rezultata analiza dr. Vlašića u desetgodišnjem razdoblju (tablica 1,2,3) vidljivo je da postotak funkcionalno muških cvjetova varira u pojedinim godinama: kod lastovke od 17 % (1956.) do 46 % (1968.); oblice od 24 % (1963.) do 72 % (1964.), a kod dužice od 18 % (1967.) do 75 % (1963.). Vidljivo je da su razlike u postotku muških cvjetova u pojedinim godinama velike. Potrebno je istaknuti da je ta razlika veća ili manja ovisno o sorti. Taj postotak i razlika veći su kod oblice i dužice, a manji kod lastovke. Prema tome, tu je prisutan utjecaj sorte, odnosno genetska komponenta.
Tablica 1. Lastovka - odnos broja cvjetova u cvatu i hermafroditnih cvjetova
Godina
Broj cvjetova u cvatu
Broj cvjetova - hermafroditnih
% cvjetova - herafroditnih
1954.
13,073
8,950
68,604
1955.
13,033
8,157
62,765
1956.
12,190
10,140
83,223
1957.
14,637
8,617
58,127
1958.
15,433
12,960
83,151
1962.
16,957
13,737
81,183
1963.
13,887
11,392
82,466
1964.
15,360
8,760
57,480
1965.
14,027
10,593
75,727
1966.
15,000
11,597
75,122
1967.
13,213
12,427
94,678
1968.
12,160
6,680
54,114
1969.
14,040
8,750
62,452
1970.
14,997
10,717
71,691
X
14,165
10,243
72,482
Kod proučavanja utjecaja vanjskih čimbenika na pojavu funkcionalno muških cvjetova, jedna grupa autora punu je pažnju obratila na utjecaj temperature i vlage u određeno doba godine. Treba istaknuti da su oni proučavali utjecaj temperature na sortama maslina u stakleniku, gdje su ostali čimbenici bili optimalni. Međutim, u prirodi istovremeno djeluju razni čimbenici, povoljni i nepovoljni.
Tablica 2. Oblica - Odnos broja cvjetova u cvatu i hermafroditnih cvjetova
Godina
Broj cvjetova u cvatu
Broj cvjetova - hermafroditnih
% cvjetova - herafroditnih
1954.
20,590
7,043
34,424
1955.
19,617
12,970
66,228
1956.
14,693
8,073
54,139
1957.
17,520
8,027
45,143
1958.
16,183
9,723
60,132
1961.
20,683
14,043
67,185
1962.
20,000
9,350
46,150
1963.
15,697
12,033
76,103
1964.
22,747
6,507
28,138
1965.
18,037
11,950
66,456
1966.
18,383
10,320
56,255
1967.
19,080
8,213
43,860
1968.
18,787
5,733
30,969
1969.
19,760
8,587
43,902
1970.
15,900
9,730
61,310
x
18,512
9,487
51,459
Te analize utjecaja temperaturnih prilika u siječnju, veljači i ožujku na intenzitet funkcionalno muških cvjetova pokazale su da su temperature u to doba (7 °C i niže po Hartmannu i Hoffmannu) utjecale na smanjenje funkcionalno muških cvjetova samo u pojedinim godinama i kod nekih sorata. Tako je temperatura 1963. utjecala na smanjenje broja funkcionalno muških cvjetova kod lastovke, oblice i dužice. Međutim, 1954. nije utjecala na smanjenje funkcionalno muških cvjetova. 1967. povoljno je utjecala samo kod lastovke i dužice, a nije kod oblice. Obrnuto, temperatura iznad 7 °C u spomenutom vremenskom razdoblju 1955. i 1957. rezultirala je povećanim brojem funkcionalno muških cvjetova sorata masline. To se nije ponovilo 1958. niti 1961., kada je broj funkcionalno muških cvjetova bio veliki samo kod dužice.
Tablica 3. Dužica - Odnos broja cvjetova u cvatu i hermafroditnih cvjetova
Godina
Broj cvjetova u cvatu
Broj cvjetova - hermafroditnih
% cvjetova - herafroditnih
1960.
16,100
9,133
56,117
1961.
21,067
7,020
33,679
1962.
25,257
13,090
51,209
1963.
10,960
2,693
24,626
1964.
14,213
7,633
50,265
1965.
17,893
7,367
41,309
1966.
16,613
13,280
79,156
1967.
14,147
11,733
82,132
1968.
12,920
9,867
76,478
1969.
18,880
10,960
58,960
1970.
14,947
8,387
56,167
x
16,727
9,197
54,164
Utjecaj pomanjkanja vlage u tlu na pojavu funkcionalno muških cvjetova kod naših sorata masline vode do istog zaključka. Spomenuti autori su utvrdili da pomanjkanje vlage u ožujku, travnju i svibnju uvjetuju zastoj u rastu tučka, zbog čega on atrofira pa je intenzitet pojave funkcionalno muških cvjetova velik. Ta analiza je pokazala da je u razdoblju od 1954. do 1970. manjak vlage bio sljedećih godina: 1955., 1957., 1961., 1962., 1964. i 1968. Veliki postotak funkcionalno muških cvjetova, koji se može pripisati utjecaju pomanjkanja vlage, bio je samo 1964. Kod sve tri sorte, 1968. samo kod dvije sorte, a 1961. samo kod jedne sorte.
Prema tome, rezultati proučavanja pojave morfološke sterilnosti dr. Vlašića, koja nastaje zbog negativnog utjecaja vanjskih čimbenika, ne ide u prilog tvrdnji drugih autora (Hartmann, Uriu, Spigel i dr.), prema kojima je taj oblik sterilnosti rezultat negativnog utjecaja isključivo ekoloških čimbenika. Analiza je pokazala da ni temperatura ni vlaga nisu isključivi uzročnici morfološke sterilnosti u određenoj godini.
Analiza ne bi bila kompletna ako se ne spomene utjecaj uroda pojedinog stabla na pojavu funkcionalno muških cvjetova, na ukupan broj cvjetova u cvatu i na građu samog cvata u idućoj godini.
Poznato je da od zametanja plodova pa do berbe maslina treba proći gotovo šest mjeseci (od svibnja do studenog), od koja su prva tri mjeseca vrlo sušna. U uvjetima, gdje se maslina ne navodnjava, nakon godine dobrog uroda, nastupa godina s visokim brojem funkcionalno muških cvjetova.
Rezultati proučavanja pojave morfološke sterilnosti lastovke, oblice i dužice pokazuju da ta sterilnost cvjetova nije stalna i nepromjenljiva veličina, već je varijabilna rezultanta djelovanja internih i ekoloških čimbenika. Na taj način rezultati proučavanja te pojave potvrđuju mišljenje treće grupe autora (Brooks, Pastore, Francini i dr.). Morfološka sterilnost izražena u pojavi funkcionalno muških cvjetova sortna je karakteristika pa se u tom pogledu naše sorte lastovka, oblica i dužica bitno razlikuju. Međutim, intenzitet te pojave varijabilna je veličina, koja ovisi o vrlo kompleksnom djelovanju i genetskih i ekoloških čimbenika.
Rezultati, do kojih je došao dr. Vlašić proučavanjem morfološke sterilnosti sorata masline, predstavljaju vrijedna saznanja, koja unose malo svjetla u problem nerodnosti stabala masline. Nužno je bilo posebnu pažnju obratiti mikrospori lastovke, oblice i dužice, to više što u literaturi nema podataka o proučavanju osobina polenovog zrna sorata masline.
Neki autori razlikuju samo normalna i sitna polenova zrna. Tom je analizom utvrdđeno da kod sorata masline postoje razni oblici polenovih zrna: okrugli, jajoliki, trokutasti i četvrtasti. Kod lastovke i oblice više je zastupljen trokutasti oblik (51 i 58 %), kod dužice više ima okruglog oblika (48 %), a manje trokutastog (35 %). Četvrtasti oblik pojavljuje se samo kod oblice i to samo 8,2 % te sasvim malo kod dužice (0,88 %), a kod lastovke nije pronađen.
Treba istaknuti da najveće dimenzije, kod sve tri proučavane sorte, imaju trokutasta polenova zrnca (četvrtasti oblik kod oblice i dužice može se zanemariti). Zatim, po krupnoći na drugo mjesto dolazi jajoliki oblik, a najsitnija su zrna okruglog oblika. Može se konstatirati da su dimenzije polenovog zrna općenito različite, ovisno o sorti, a unutar svake sorte veličina polenovog zrna su različita, ovisno o njihovom obliku.
Ta razmatranja dovode do konstatacije da je veličina polenovog zrna funkcija njegovog oblika, zato je oblik polenovog zrna u uskoj vezi s njegovom funkcionalnošću. Potvrda toj konstataciji su i rezultati klijavosti polena te citokemijske osobine polena.
Vrijedno je citirati rezultate proučavanja morfoloških svojstava polena sorata jabuke, koje je izvršila Taranova (1968.), koja je utvrdila da su kod sorata, koja imaju polenova zrna homogene veličine, ta zrna pretežno trokutastog oblika. Naprotiv, sorte, kod kojih su polenova zrna heterogenih veličina, imaju više zrna okruglog i jajolikog oblika.
Kod istraživanih sorata masline stanje je sljedeće: kod lastovke zrna trokutastog oblika čine 51 %, kod oblice 59 %, a kod dužice samo 35 %. Ostalo su polenova zrna drugih oblika i veličina. Kod dužice polenova zrna su jako heterogena po veličini i obliku. Ona su heterogena i kod lastovke i oblice, ali je ipak nešto povoljnija situacija. Ako rezultate analize kompariramo s rezultatima klijavosti polena, onda se može utvrditi da je i oblik, prema tome i homogenost oblika, u pozitivnoj korelaciji s fertilitetom polenovog zrna. Iz toga se može izvući još jedna konstatacija, neujednačenost oblika polenovog zrna govori da je proces formiranja mikrospore u raznim fazama tekao različito. Zato su polenova zrna tih sorata više ili manje morfološki neujednačena.
Konstataciju, da normalan proces formiranja mikrospore masline dovodi do trokutastog oblika, potvrđuje i analiza broja pora u polenovom zrnu. Općenito, kod lastovke, oblice i dužice najviše ima polenovih zrna s tri pore. Međutim, postotak polenovih zrna s tri pore znatno je veći od postotka trokutastog oblika kod sve tri sorte. Ta činjenica govori da tri pore imaju, ne samo trokutasta polenova zrna, već i jedan dio okruglih zrna. Neujednačenost u obliku i veličini uvjetuje nisku klijavost i smanjuje funkcionalnu sposobnost polenovog zrna.
U ispitivanjima intenziteta klijavosti polena sve tri sorte utvrđeno je da je najpovoljnija koncentracija saharoze kod oblice 12, 14, i 16 %, a kod lastovke i dužice 14 %. Pri toj koncentraciji saharoze, nakon šest sati, postotak klijavosti je vrlo malen i iznosi kod dužice svega 14,42 %, kod lastovke 25,53 %, a kod oblice 34,81 %. Kako je vidljivo, jedino oblica ima klijavost polena malo iznad 30 % pa se prema klasifikaciji „Florina“ ubraja u sorte dobre klijavosti, dok su lastovka i naročito dužica, sorte sa slabom klijavosti polena. Treba istaknuti da klijavost polena znatno varira od godine do godine, tako da kod oblice varira ispod 30 %, odnosno kod lastovke iznad 30 %. Općenito, klijavost polena je slaba kod lastovke i oblice, a vrlo slaba kod dužice. Agati je još 1951. utvrdio vrlo slabu klijavost polena sorte frantoio, svega 18,8 %, dok je Almeida, kod sorata masline u Portugalu, ustanovio još manju klijavost polena i to svega 0,8 - 12 %. Rezultati dr. Vlašića su, prema tome, u skladu s rezultatima, koje su postigli i drugi autori.
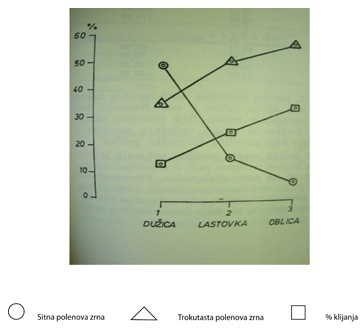
Grafikon 1. Odnos između % klijanja, sitnih i trokutastih polenovih zrna
Rezultati intenziteta klijavosti polena u skladu su s analizom morfoloških osobina polenovog zrna. Ustanovljeno je da su polenova zrna vrlo neujednačena, naročito kod dužice, što se i odrazilo na rezultate klijavosti polena. Da između oblika polenovog zrna i intenziteta klijanja postoji uska veza vrlo dobro ilustrira grafikon 1.
Oblica ima više trokutastih zrnaca, a manje okruglih te su njena zrna krupnija. Naprotiv, dužica ima manje trokutastih zrnaca, a puno više okruglih i sitnih te veliku neujednačenost u dimenzijama polenovog zrna. Lastovka je u tom pogledu između te dvije sorte. Zato i rezultat klijanja polena odražava takvo stanje i još jednom potvrđuje postavku da su trokutasta zrnca polena fertilna.
Ne samo oblik već i brzina klijanja i dužina polenove cjevčice odraz su također funkcionalnosti polenova zrna. Ta proučavanja pokazala su nešto veći intenzitet rasta polenove cjevčice kod lastovke, nakon 4 sata, kod oblice nakon 6 sati, dok je intenzitet rasta cjevčice kod dužice vrlo malen. Općenito se može tvrditi da je sposobnost klijanja polenove cjevčice kod lastovke, oblice i dužice malena, što je u skladu s morfološkom analizom polenovog zrna i s klijavosti polena.
Nameće se pitanje koji čimbenici uvjetuju veliku neujednačenost polenovog zrna, nisku klijavost i slabi porast polenove cjevčice. To vodi proučavanju citogenetičkih osobina, analiziu kemijskih sastojaka i analizi citokemijskih osobina polena tih sorata.
Kod proučavanja procesa mikrosporogeneze posebna se pažnja daje pojavama abnormalnosti, koje se javljaju u metafazi i telofazi te kod asinhronog tijeka diobe i abnormalnosti u fazi tetrada.
Pojava nepotpune kongresije kromosoma utvrđena je u metafazi i telofazi. Zakašnjeli kromosomi imaju strukturu bivalenata, iako ima pojedinih slučajeva kada nije jasno da li se radi o bivalentima ili o univalentima. Ta se opažanja slažu s opažanjima nekih talijanskih autora, koji također pretpostavljaju da se u rijetkim slučajevima radi o univalentima u zakašnjenju. Kod zakašnjelih kromosoma u metafazi i telofazi postoji razlika između sorata, između pojedinih faza te to varira od godine do godine. U prosjeku, intenzitet te abnormalnosti iznosi kod lastovke 16 %, kod oblice 17 %, a kod dužice oko 19 %.
Druga abnormalnost je asinhroni tijek diobe stanice u istoj anteri. Proces diobe traje vrlo kratko pa postoji mogućnost da neka stanica u zakašnjenju dovede do kraja samo tijek diobe, ali može degenerirati stanica, koja kasni u procesu diobe.
Na temelju proučavanja te pojave utvrđeno je da asinhroni tijek diobe stanice kod lastovke, oblice i dužice nije čest.
Detaljna analiza pojave abnormalnosti u fazi tetrada omogućila je da se utvrdi ne samo pojava već i njezin intenzitet. U toj fazi utvrđeno je prisustvo, pored normalnih tetrada, monada, diada, trijada, abnormalnih tetrada, pentada i heksada. Od abnormalnih, u toj fazi najviše su zastupljene tetrade, trijade i pentade. Međutim, u intenzitetu tih abnormalnosti postoje razlike između sorata kao i između godina. U prosjeku one iznosi kod lastovke 14%, a kod dužice i oblice oko 15%. Nakon analize ovih rezultata ispitivanja abnormalnosti u procesu mikrosporogeneze možemo konstatirati da kod lastovke, oblice i dužice ima abnormalnosti u ovom procesu.
Međutim, analiza pojave abnormalnosti u procesu mikrosporogeneze, mada daje korisne podatke (kao što i sam autor kaže), ne rasvjetljava u potpunosti problem vrlo niske klijavosti polenovog zrna naših sorata masline.
Sadržaj slobodnih saharida, prema tim analizama, je različit, ovisno o sorti. Polen lastovke sadrži najviše saharida (2,75 %), najmanje ih sadrži polen dužice (1,57 %). Ti rezultati potvrđuju rezultate drugih autora, prema kojima sadržaj šećera ovisi o vrsti i o sorti te su u skladu s rezultatima drugih autora, koji tvrde da morfološki ujednačeni polen ima veći sadržaj šećera. Kod analize morfologije polena praćenih sorata utvrđena je neujednačenost polena, naročito kod dužice. Ta sorta ima i najmanju količinu slobodnih saharida.
Na temelju analize sadržaja slobodnih saharida kod lastovke, oblice i dužice, pored osjetne razlike u sadržaju glukoze i fruktoze između sorata, može se prijetiti da su količine glukoze, a naročito fruktoze vrlo niske, što se negativno odražava na metabolizam polenova zrna.
Analitički podaci za aminokiseline u polenu ispitanih sorata pokazali su razliku kvalitete i kvantitete proteinskog sastava. U polenu ispitivanih sorata masline utvrđeno je 16 aminokiselina.
U pogledu ukupnih količina slobodnih aminokiselina između sorata masline nema izrazite razlike, one su vidljive u sadržaju pojedinih aminokiselina. Tako polen lastovke sadrži više prolina, glicina, alanina, asparagina i metionina od ostalih sorata. Tu se ističe važnost, koju ima za izgradnju fiziološki aktivnog polena, asparagin, metionin, a za klijavost naročito prolin. U polenu oblice ima više slobodnog treonina i histidina, dok u polenu dužice ima više ostalih aminokiselina u slobodnom obliku. Razlika u kvantiteti pojedinih slobodnih aminokiselina govori o različitoj kvaliteti polenova zrna lastovke, oblice i dužice. Treba istaknuti da sadržaj slobodnih aminokiselina označava potencijalnu sposobnost za sintezu biokemijskih sastojaka pa prema tome i za odvijanje biokemijskih procesa, o kojima ovisi redovita funkcionalnost polena. Količine pojedinih aminokiselina u maslini su vrlo male, ako ih se usporedi s količinama aminokiselina u polenu nekih voćnih vrsta.
Pored analize sadržaja saharida i aminokiselina u polenu, za funkcionalnost samog polenovog zrna od bitne važnosti su i nukleinske kiseline. Rezultati analize sadržaja histona u polenu lastovke, oblice i dužice, pokazuju prije svega razliku između sorata. Veći sadržaj histona utvrđen je u polenu lastovke, zatim u polenu dužice. Međutim, važno je analizirati razliku u sadržaju histona između raznih oblika polenova zrna. U tom pogledu trokutasta zrna lastovke, odnosno trokutasta i četvrtasta zrna oblice, imaju više histona od jajolikih i okruglih zrna. Ta je razlika naročito izražena kod lastovke, kod oblice je nešto manja. Kod dužice trokutasta polenova zrna imaju manje ekstinkcione vrijednosti (izumiranje, propadanje), a veće imaju okrugla zrna.
Neujednačenost histonskog sadržaja u raznim oblicima polena mora imati određenog utjecaj i na sadržaj nukleinske kiseline pa, prema tome, i na klijavost polena i rast polenove cjevčice. Ekstinkcione vrijednosti za histone, na temelju tih analiza, vrlo su niske. Kod lastovke čine 47 i 41%, kod oblice 56 i 59 %, a kod dužice 54 i 52 % od optimalnog iznosa.
Analiza sadržaja DNA utvrdila je znatnu razliku između pojedinih oblika polenova zrna. Najveći sadržaj DNA ustanovljen je su trokutastim i četvrtastim oblicima polenova zrna kod lastovke i oblice, kod dužice razlike između oblika su znatno manje. Tim je analizama ustanovljen različit intenzitet neujednačenosti polenovog zrna, od sitnih do jako krupnih mikrospora. Prema tome, normalno je očekivati i različite vrijednosti DNA u polenu različitog oblika i veličine, što je analizama i utvrđeno.
Sadržaj RNA u polenu je različit, u ovisnosti o obliku polenova zrna. Odnos između DNA i RNA ovisi o veličini jezgre i citoplazme pa u stanicama, koje sadrže više DNA, ima manje RNA i obrnuto. U polenu ispitivanih sorata masline ima općenito više DNA kod svih oblika, prema tome može se reći da su svi oblici polenovih zrna bogatiji sadržajem jezgre nego citoplazme.
Smatra se da je niski sadržaj histona i nukleinskih kiselina kod ispitivanih sorata masline, u odnosu na neke druge voćne vrste, odraz slabe funkcionalnosti polenova zrna.
U tom radu utvrđene su razlike u ekstinkcionim vrijednostima za histone, DNA, RNA i ukupne nukleinske kiseline, za okrugli i jajoliki oblik polenova zrna jako su izražene između lastovke, oblice i dužice. Razlike u citokemijskim svojstvima kod trokutastog oblika su vrlo male i vrijednosti su skoro iste.
Prema tome, rezultati analiza dovode do konstatacije da trokutasti oblik polenova zrna predstavlja normalno izgrađeni i funkcionalni oblik, dok su nepotpuno i nedovoljno izgrađeni ostali oblici pa su zato i defektni.
Ujednačenost citokemijskih osobina kod trokutastog oblika polenova zrna govori da između lastovke, oblice i dužice ne postoji razlika. Razlike su, međutim, jako izražene kod okruglog i jajolikog oblika. Dakle, velika neujednačenost u oblicima polenova zrna odraz je funkcionalnosti, odnosno slabe klijavosti polena. Velika neujednačenost u oblicima polena, naročito kod dužice, odnosno mala zastupljenost trokutastog oblika, uvjetuje nisku klijavost polena kod naših sorata masline.
Kod njih je česta pojava nedovoljnog zametanja plodova, mada je broj hermafroditnih cvjetova bio visok. Treba imati na umu da je maslina anemofilna biljka i da se oprašivanje vrši putem laganog vjetra. Zato uspješno oprašivanje i dobro zametanje plodova ovisi o mnogo čimbenika.
Ovim desetogodišnjim ispitivanjem fenofaze cvjetanja kod lastovke, oblice i dužice utvrđeno je da početak te fenofaze kao i njezino trajanje ovisi o temperaturnim i oborinskim prilikama, tijekom ožujka, travnja i svibnja. Niske temperature i mnogo oborina u tom razdoblju uvjetuju kašnjenje te fenofaze i obrnuto. Dužina fenofaze varira od 10 do 13 dana.
Ova proučavanja uzela su u obzir i intenzitet otvaranja cvjetova sorata masline. Najveći broj cvjetova ispitivanih sorata otvara se u vrlo kratkom roku, svega 4 - 5 dana. Treba još uzeti u obzir da su cvjetovi, koji se kasnije otvaraju, u većem postotku funkcionalno muški pa se iz toga može vidjeti kako je vrijeme, u kojem se treba izvršiti oprašivanje, vrlo kratko. Međutim, pri tom je važno utvrditi i dužinu vitalnosti embrionalne kese.
O građi embrionalne kese postoje različita tumačenja. Postoji tvrdnja da ima četiri jezgre i da se razvija po tipu Endymion, druga tvrdnja je da ima osam jezgri i da se razvija po tipu Allium. Tom analizom utvrđeno je da embrionalna kesa ima, pored jajne stanice, sinergide, sekundarne jezgre i antipode. Ti rezultati ističu da embrionalna kesa kod maslina ima osam jezgri. Međutim, činjenica je da su antipode vidljive vrlo kratko. Zbog toga su, vjerojatno, neke ranije tvrdnje bile da embrionalna kesa ima četiri jezgre.
Neki su pak autori proučili utjecaj, koji ima temperatura, na brzinu rasta polenove cjevčice nakon oprašivanja vlastitim i stranim polenom. Utvrdili su da je i kod povoljnih temperatura rast polenove cjevčice vrlo spor pa ako i dođe do embrionalne kese, ne može oploditi jajnu stanicu, jer ona degenerira ranije.
Na temelju rezultata tih pokusa utvrđeno je da je dužina vitalnosti embrionalne kese sortna osobina. Kod oblice je ona vrlo kratka, dok je kod lastovke i dužice duža. Rezultati istraživanja pokazuju veliku praktičnu vrijednost i u skladu su s opažanjima o urodu tih sorata masline u pojedinim godinama.
Međutim, za uspjeh oprašivanja i zametanja plodova presudnu ulogu ima kompatibilitet, bilo da se radi o auto ili interkompatibilitetu.
Na temelju rezultata pokusa o intenzitetu samooplodnje, utvrđeno je da uspjeh samooplodnje znatno varira od godine do godine i da je rezultat djelovanja i internih i vanjskih čimbenika. Tako variranje intenziteta samooplodnje iznosi kod lastovke od 0,002 % do 0,44 %, kod oblice 0,07 % do 1,22 %, a kod dužice 0,04 % do 0,89 % i različito je, ovisno o sorti.
Prema tvrdnji nekih autora, sorte maslina su autoinkompatibilne, ako uspjeh oplodnje na temelju samooprašivanja iznosi 1 - 2 ‰. U tom pogledu naše su sorte u nekim godinama autoinkompatibilne. Rezultat pokusa potvrđuje da su sorte lastovka, oblica i dužica djelomično samooplodne i praktički autosterilne.
Kao što su utvrdili pojedini autori, kod sorata, kod kojih je utvrđen autoinkompatibilitet u većem ili manjem intenzitetu, treba očekivati i pojavu interinkompatibiliteta, kojima su uzroci iste naravi.
Rezultati proučavanja pojave interinkompatibiliteta potvrdili su tu konstataciju. Veći stupanj interkompatibiliteta s lastovkom, oblicom i dužicom imaju sorte drobnica, levantinka, uljarica i piculja, a manji stupanj kompatibiliteta imaju ostale sorte.
Ipak, treba istaknuti da taj stupanj kompatibiliteta izražen u postotku oplodnje varira od godine do godine, zbog djelovanja i utjecaja mnogih čimbenika. U tom se naročito ističe dužica, kod koje je razlika u pojedinim godinama znatna. Općenito se može tvrditi da lastovka pokazuje veći stupanj interkompatibiliteta s prvom grupom sorata, nego oblica i dužica, jer je postotak oplodnje s tim sortama veći. Prema tome, na temelju tih pokusa i analize podudarnosti u fenofazi cvatnje, dobri oprašivači za oblicu i dužicu jesu sorte drobnica, levantinka i uljarica, a za lastovku se može preporučiti samo sorta piculja, jer ostale kompatibilne sorte cvjetaju kasnije.